最危险的沙粒病毒 拉沙病毒
最危险的沙粒病毒 拉沙病毒

杜鹃
杜鹃:今天介绍的是拉沙病毒(Lassa Mammarenavirus/Lassa Virus/LASV)
简介
拉沙病毒(LASV)是一种沙粒病毒,会在人和其他灵长类动物中引起拉沙出血热,是一种病毒性出血热(VHF)的元凶。 拉沙病毒是一种新兴病毒,需要生物安全等级为4的等效收容措施。 它在西非国家特别流行,特别是塞拉利昂,几内亚共和国,尼日利亚和利比里亚,那里的年感染率在300000至500000例之间,每年导致5000例死亡。
截至2012年,西非马诺河地区的发现扩大了两个已知的拉沙病毒流行地区之间的流行区,这表明LASV在整个西非热带树木稀树草原生态区中分布更为广泛。没有批准用于人类的拉沙病毒疫苗。
发现
1969年,传教护士劳拉·怀恩(Laura Wine)在尼日利亚博尔诺州的一个村庄拉萨(Lassa),一名产科患者患上了一种神秘疾病,她因此染上了病。然后,她被运送到尼日利亚的乔斯,并在那里去世。 随后,另外两个被感染,其中一个是五十二岁的护士莉莉·皮尼奥(Lily Pinneo),她曾照顾劳拉·怀恩(Laura Wine)来自Pinneo的样本被送至纽黑文的耶鲁大学,在那里,Jordi Casals,Sonja Buckley等人首次分离出一种新病毒,后来被称为拉沙病毒。卡萨尔斯发烧,几乎丧命。 一名技术人员因此丧命。到1972年,多乳鼠Mastomys natalensis被发现是西非拉沙病毒的主要储存库,能够在尿液和粪便中排出病毒而没有明显的症状。
病毒特征
拉沙病毒
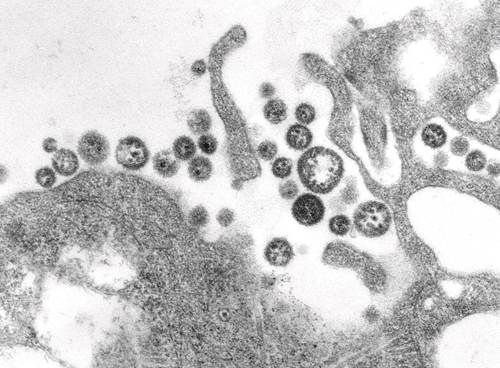
电子显微镜下的拉沙病毒
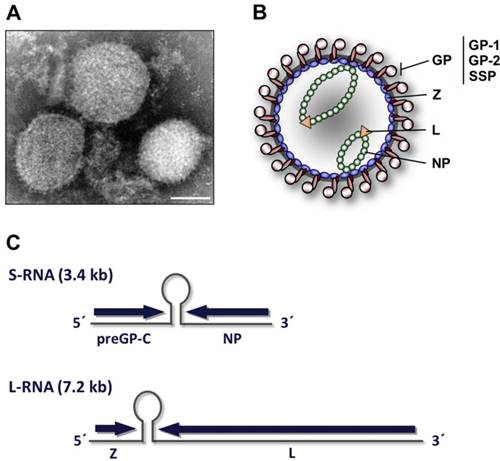
拉沙病毒是有包膜的,单链的,双节段的,双义性的RNA病毒。 它们的基因组包含在两个RNA片段中,每个RNA片段编码两种蛋白质,每种意义上一种,共编码四种病毒蛋白质。大片段编码一个小的锌指蛋白(Z),它调节转录和复制和RNA聚合酶(L)。 小片段编码核蛋白(NP)和表面糖蛋白前体(GP,也称为病毒刺突),通过蛋白水解切割成包膜糖蛋白GP1和GP2,后者与α-dystroglycan受体结合并介导宿主细胞进入。
拉沙病毒引起拉沙出血热,通常通过免疫抑制表现出来。 拉沙病毒复制速度非常快,并且在复制中表现出时间控制。第一个复制步骤是转录反义或反义基因组的mRNA拷贝。 由于NP和L蛋白是从mRNA翻译而来的,因此可以确保为后续的复制步骤提供足够的病毒蛋白。 然后,正义或正义基因组会复制自身的病毒互补RNA(vcRNA)。RNA拷贝是产生反义后代的模板,但也可以从中合成mRNA。 由vcRNA合成的mRNA被翻译成GP和Z蛋白。 这种时间控制使得刺突蛋白最后产生,因此,延迟了宿主免疫系统的识别。
对基因组的核苷酸研究表明,拉沙病毒有四个血统:三个在尼日利亚,四个在几内亚,利比里亚和塞拉利昂。 尼日利亚菌株似乎是其他菌株的祖先,但需要进一步的工作来证实这一点。
病毒受体
拉沙病毒入侵
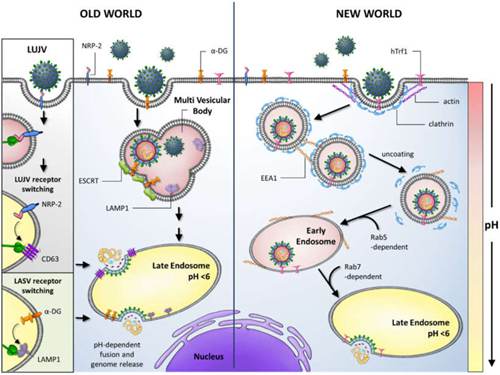
拉沙病毒通过细胞表面受体α-dystroglycan(alpha-DG)进入宿主细胞,α-dystroglycan(21-DG)是细胞外基质蛋白的多功能受体。 它与原型旧世界性沙粒病毒的淋巴细胞性脉络膜脑膜炎病毒共享该受体。 受体识别取决于被称为大蛋白的一组糖基转移酶对α-dystroglycan的特定糖修饰。 在拉沙病毒流行的西非,编码这些蛋白质的基因的特定变体似乎处于正选择状态。α-肌营养不良糖还被新大陆进化枝C沙粒病毒(Oliveros和Latino病毒)的病毒用作受体。 相比之下,进化枝A和B的新世界舞台病毒,除了非致病性阿马帕里病毒(Amapari virus)外,还包括重要的Machupo病毒、Guanatoto病毒、Junin病毒和Sabia病毒,还使用了转铁蛋白受体1。GP1糖蛋白上的一个小脂肪族氨基酸 氨基酸位置260是与α-DG高亲和力结合所必需的。 另外,GP1氨基酸259位似乎也很重要,因为所有显示出高亲和力的α-DG结合的沙粒病毒都在此位置具有庞大的芳香族氨基酸(酪氨酸或苯丙氨酸)。
与大多数包膜病毒使用网格蛋白包被的凹坑进行细胞进入并以pH依赖方式结合其受体不同,拉沙病毒和淋巴细胞性脉络膜脑膜炎病毒使用的胞吞途径独立于网格蛋白、小窝蛋白、动力蛋白和肌动蛋白。 一旦进入细胞内,病毒便会通过囊泡运输迅速传递至内体,尽管这种病毒在很大程度上独立于小的GTPases Rab5和Rab7。 与内体接触时,pH依赖性膜融合发生在包膜糖蛋白的介导下,该蛋白在内体的较低pH下与溶酶体蛋白LAMP1结合,从而导致膜融合并从内体逃逸。
病毒生活史
拉沙病毒生活史
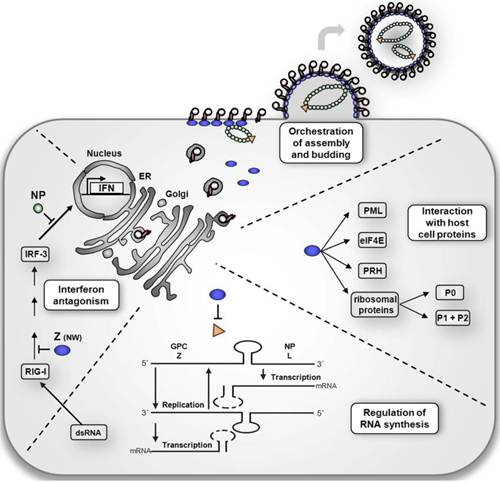
拉沙病毒的生命周期与旧世界的鼻病毒相似。 拉沙病毒通过受体介导的内吞作用进入细胞。 尚不清楚使用哪种胞吞途径,但至少细胞进入对胆固醇消耗敏感。 据报道,病毒的内在化受胆固醇消耗的限制。 用于细胞进入的受体是α-dystroglycan,这是一种高度保守且普遍存在的细胞外基质蛋白的细胞表面受体。dystroglycan,后来被切割成α-dystroglycan和β-dystroglycan,最初在大多数细胞中表达到成熟组织,并且它在ECM和基于肌动蛋白的细胞骨架之间提供了分子联系。病毒通过α-dystroglycan介导的内吞作用进入细胞后,低pH环境触发pH依赖性膜融合并将RNP(病毒核糖核蛋白)复合物释放到细胞质中。病毒RNA被解压缩,并在细胞质中开始复制和转录。随着复制的开始,S和L RNA基因组都合成了反基因组S和L RNA,并从反基因组RNA中合成了基因组S和L RNA。 基因组和反基因组RNA都是转录和翻译所必需的。S RNA编码GP和NP(病毒核衣壳蛋白)蛋白,而L RNA编码Z和L蛋白。L蛋白最有可能代表病毒依赖RNA的RNA聚合酶。当细胞被病毒感染时,L聚合酶与病毒RNP结合并启动基因组RNA的转录。 两个RNA片段的5和3末端19 nt病毒启动子区域对于识别和结合病毒聚合酶都是必需的。 初级转录首先转录来自基因组S和L RNA的mRNA,分别编码NP和L蛋白。 转录终止于基因组区域内的茎环(SL)结构。 沙眼病毒使用帽抢夺策略从细胞mRNA中获得帽结构,并且它是由L聚合酶的核酸内切酶活性和NP的帽结合活性介导的。 抗基因组RNA分别从S和L片段转录以基因组方向编码的病毒基因GPC和Z。 反基因组RNA也是复制的模板。GPC翻译后,在内质网中进行了翻译后修饰。GPC在分泌途径的后期分裂为GP1和GP2。 据报道,细胞蛋白酶SKI-1 / S1P负责这种切割。 当病毒发芽并从细胞膜释放时,切割的糖蛋白被掺入病毒体膜中。
相关疾病
拉沙热主要是由拉沙病毒引起的。 症状包括以发烧,全身无力,咳嗽,喉咙痛,头痛和胃肠道表现为特征的流感样疾病。 出血表现包括血管通透性。
进入后,拉萨病毒会感染人体的几乎所有组织。 它从粘膜,肠,肺和泌尿系统开始,然后发展到血管系统。
病毒的主要靶标是抗原呈递细胞,主要是树突状细胞和内皮细胞。在2012年,有报道说拉沙病毒核蛋白(NP)会破坏宿主的先天免疫系统反应。 通常,当病原体进入宿主时,先天防御系统会识别与病原体相关的分子模式(PAMP)并激活免疫反应。 其中一种机制是检测仅由反义病毒合成的双链RNA(dsRNA)。 在细胞质中,dsRNA受体[例如RIG-1(视黄酸诱导基因I)]和MDA-5(黑色素瘤分化相关基因5)检测dsRNA并启动易位IRF-3(干扰素调节因子3)和其他转录因子进入细胞核。 易位的转录因子激活干扰素α的和表达,并启动适应性免疫。 拉萨乳病毒中编码的NP在病毒复制和转录中必不可少,但它也通过抑制IRF-3的易位而抑制宿主固有的IFN反应。 据报道,拉沙病毒的NP仅对dsRNA具有核酸外切酶活性。NP dsRNA核酸外切酶活性可通过消化PAMP来抵消IFN的应答,从而使病毒能够逃避宿主的免疫应答。
(最危险的沙粒病毒 拉沙病毒)宝,都看到这里了你确定不收藏一下??